
Seroprevalence and Risk Factors for Toxoplasma gondii and Neospora caninum in Cattle in Portugal


1
Laboratório de Parasitologia, Instituto Nacional de Investigação Agrária e Veterinária, Av. da República, Quinta do Marquês, 2780-157 Oeiras, Portugal
2
Centre for Interdisciplinary Research in Animal Health (CIISA), Faculty of Veterinary Medicine, University of Lisbon, 1300-477 Lisboa, Portugal
3
Associate Laboratory for Animal and Veterinary Sciences (AL4AnimalS), 1300-477 Lisbon, Portugal
4
Friedrich-Loeffler-Institute, Institute of Epidemiology, 17493 Greifswald, Insel Riems, Germany
*
Author to whom correspondence should be addressed.
Animals 2022, 12(16), 2080; https://doi.org/10.3390/ani12162080
Submission received: 31 May 2022
/
Revised: 4 August 2022
/
Accepted: 6 August 2022
/
Published: 15 August 2022
(This article belongs to the Special Issue Serological and Molecular Epidemiology in Animals)
Abstract
:Simple Summary
Neospora caninum and Toxoplasma gondii are apicomplexan parasites with a worldwide distribution and both infect cattle. While the importance of N. caninum is mainly linked to reproductive losses, concern has been raised about the role of beef as a source of food-borne toxoplasmosis in humans. Little is known about the prevalence of these parasites in cattle in Portugal. This study aimed to evaluate the seroprevalence and risk factors for N. caninum and T. gondii in naturally exposed cattle from different geographical areas in the country. Results point to a low but widespread seroprevalence of T. gondii, while that of N. caninum was found to be in the same range as reported in other Mediterranean countries. Distribution of both parasites may be explained by common climate factors and management practices identified as significant predictors of infection in the study.
Abstract
Neospora caninum has a worldwide economic impact as an important cause of abortion in cattle, while Toxoplasma gondii, another abortifacient pathogen, is globally a major foodborne zoonotic threat. The study aimed to evaluate the seroprevalence and risk factors for the two parasites in cattle in Portugal. A total of 612 sera from 35 farms were tested by an in-house p30 ELISA for T. gondii and p38 ELISA for N. caninum. T. gondii positive and suspicious sera were confirmed by p30 Western blot or IFAT. T. gondii and N. caninum animal seroprevalence was 9.2% (95%CI 7.1–11.7) and 17.2% (95% CI 14.4–20.4) and herd seroprevalence was 51.4% (95% CI 35.6–67.0) and 68.6% (95% CI 52.0–81.5), respectively. At the univariable level, climate area and precipitation of wettest month, driest month, driest quarter, and warmest quarter were significant predictors of seropositivity for both. N. caninum seropositivity was more likely in the region Norte, densely populated areas, and intensive production, and the probability of T. gondii seropositivity decreased with herd size. Results confirm the need to consider neosporosis in the differential diagnosis of cattle reproductive disorders in Portugal and may be valuable to inform source attribution models for human toxoplasmosis.
1. Introduction
Neospora caninum and Toxoplasma gondii are obligate intracellular cyst-forming coccidian parasites with a worldwide distribution. Both parasites are capable of infecting a wide range of warm-blooded hosts. N. caninum is an important cause of abortion in cattle [1] with a considerable impact on the beef and dairy industries. Economic losses arise mainly from the lower reproductive performance of cattle [1,2] but were also linked to lower milk production, a decrease in weight gain, and premature culling [2,3,4]. T. gondii is recognized as a common cause of abortion in small ruminants but does not seem to play a relevant role in reproductive disorders of cattle [5]. Unlike N. caninum, whose zoonotic potential has never been clearly elucidated, T. gondii is highly ranked among foodborne pathogens. Human toxoplasmosis is particularly dangerous in its congenital form and in people with immunological deficiency. Furthermore, in immunocompetent individuals, where infection is generally considered asymptomatic, ocular toxoplasmosis is currently more often associated with acquired infection than with congenital infection [6,7]. In addition, there has been a growing interest on the influence of T. gondii on cognition and mental disorders [8]. Both parasites have a typical heteroxenous apicomplexan lifecycle, with canids and felids acting as definitive hosts for N. caninum and T. gondii, respectively. Infection of cattle occurs through the ingestion of oocysts in contaminated food and water, while carnivorous and omnivorous hosts may be infected either by the fecal–oral route or by the ingestion of viable tissue cysts in meat. Vertical transmission, another route of infection, may occur through multiple generations in the case of N. caninum, being considered the main mode of transmission in cattle herds, but does not seem to play an important role in the case of T. gondii. In humans, transplacental infection may occur in non-immune women during pregnancy. In this regard, a European multicenter study estimated that 30–60% of seroconversions in pregnant women are linked to the consumption of raw or undercooked meat while only 6–7% are probably caused by soil contact [9].
T. gondii tissue cysts in raw meat can remain viable during several days at common fridge temperatures [10] and survive up to 5 min when heated at temperatures of 60 °C to 100 °C [11].
While the importance of cattle as a source of human infection is generally known [9,12], public health risks through the consumption of beef and cattle-derived products, such as artisan fresh cheese, which was already linked to an outbreak of acute toxoplasmosis in humans, remain unclear [13]. Hence, seroprevalence studies on T. gondii in cattle may provide valuable input data for quantitative source attribution models to measure the risk of human infection attributable to the consumption of beef. On the other hand, improving knowledge on N. caninum prevalence in the cattle population is important to evaluate the extent and impact of infection on a wider scale and consider this parasite in the differential diagnosis of abortive conditions. In Portugal, little information on the prevalence of T. gondii and N. caninum in cattle is available, therefore, the aim of this study was to evaluate the presence of specific antibodies against both parasites in naturally exposed animals using a panel of well-characterized sera from different geographical locations.
2. Materials and Methods
2.1. Sample Characterization
Serological tests were performed on 612 cattle sera selected from a serum collection from a previous cross-sectional study on bovine besnoitiosis in Portugal [14] available at our institute. Blood samples were obtained between 2012 and 2013 using a two-stage cluster sampling scheme, in which a random representative number of farms was selected first and then a random representative number of animals was selected per farm [14]. Sera were stored frozen in 1–1.5 mL aliquots at −20 °C in a temperature-controlled freezer connected to temperature monitoring sensors and thawed for not more than 2–3 times prior to serological testing. For the purpose of this study, a convenience sample was used in which a total of 35 of the above-mentioned farms were selected to cover as much as possible the different geographical areas in Portugal. The number of serum samples available for each farm ranged between 6 and 24. Therefore, for an estimated herd prevalence of 20%, assuming a 90% sensitive test, the number of animals tested per farm should allow identifying at least one positive animal in 32 of the farms with 95% confidence, and with 70–75% confidence in the remaining 3 farms [15]. Animal and farm data (age, sex, herd size, production system and production type, farm location) were available from the former study [14]. All sera were negative for antibodies to Besnoitia besnoiti [14].
2.2. Testing Strategy
Serological testing for N. caninum was performed using an in-house p38 ELISA [16]. Serum samples were tested for antibodies to T. gondii by a sequential testing strategy, with an in-house p30 ELISA [17,18] as the first screening assay followed by the p30 Western blot (p30WB) [18,19,20] and Immunofluorescent Antibody Test (IFAT) [21] as confirmatory tests for positive and suspicious results. There is no reference standard to assess seropositivity for T. gondii in cattle. Thus, the cut-off for seropositivity by the p30 ELISA was determined based on the empirical 95th percentile method and the cut-off for suspicious sera was set at the 90th percentile, i.e., regarding the 5% strongest reactions as positive and the remaining 10% strongest reactions as suspicious. Confirmation of negative results by the p30 WB and IFAT was performed for a panel of randomly selected negative sera (n = 31).
2.2.1. Neospora caninum p38-ELISA
The p38 ELISA was carried out as described earlier [16]. The wells of 96-well Polysorb ELISA plates (Nunc) were coated with 120 μL affinity-purified native N. caninum p38 tachyzoite surface antigen diluted in 0.1 M bicarbonate buffer (pH 8.3) to a concentration of 0.1 μg/mL and incubated for 1 h at 37 °C. Plates were washed three times and blocked with 300 μL of 20% horse serum in PBS-0.05% Tween (PBS-T) for 30 min at 37 °C. Sera were diluted 1:100 in blocking buffer and distributed in 100 μL duplicates on plates. A positive control diluted 1:80 and a negative control were included in each plate. The positive serum was no longer identical to the serum mentioned in the initial report [16] but came from the same animal. It was stored in aliquots at −20 °C until use, producing identical values as the initial positive control serum, when applied at a 1:80 dilution. After 30 min at 37 °C, plates were washed three times with PBS-T and added with 100 μL monoclonal anti-bovine IgG Clone BG-18 Biotin Conjugate (Sigma) diluted 1:2000 in PBS-T containing 1% horse serum. After 30 min at 37 °C, plates were washed three times, added with 100 μL Extravidin® Peroxidase Conjugate and incubated at 37 °C for another 30 min. The reaction was revealed for 15 min at 37 °C with 100 μL substrate solution prepared with 0.2 M sodium acetate and 0.2 M citric acid containing 100 μg/mL 3,3,5,5-tetramethylbenzidine (TMB) and 0.004% hydrogen peroxide. The colorimetric reaction was stopped with 50 μL of a 2M H2SO4 solution. Optical densities were read at 450 nm on a SunriseTM microplate reader (TECAN Deutschland GmbH, Crailsheim, Germany).
2.2.2. Toxoplasma gondii p30-ELISA
The p30-ELISA was based on previously established methods [17,18] with few modifications and an adaptation to cattle samples. The wells of 96-well Polysorb ELISA plates (Nunc) were coated with 120 μL affinity-purified native T. gondii p30 tachyzoite surface antigen diluted in 0.1 M bicarbonate buffer (pH 8.3) to a concentration of 0.1 μg/mL and incubated for 1 h at 37 °C. Plates were washed three times and blocked with 300 μL of 1% casein in PBS-0.05% Tween for 30 min at 37 °C. One positive and one negative control were included in each plate. Sera and positive and negative controls were diluted 1:100 in blocking buffer and distributed in 100 μL duplicates on plates. As positive control serum, a pool of positive sheep sera was used. Reactivity with the heterologous anti-bovine conjugate was confirmed by Western blot previously (data not shown). The negative control was a serum from a calf reared at Friedrich-Loeffler-Institute and negative for T. gondii by IFAT (titer < 1:50). After 30 min incubation at 37 °C, plates were washed three times with PBS-T and developed with 100 μL Peroxidase-conjugated AffiniPure Rabbit Anti-Bovine IgG [H+L] (Jackson ImmunoResearch Laboratories) diluted 1:4000 during 30 min at 37 °C. The colorimetric reaction was stopped and optical densities were read as described for the N. caninum p38 ELISA.
2.2.3. Western Blot
Western blot analysis to detect antibodies against T. gondii p30 protein was performed as described previously [18,19,20] but adapted to cattle samples. Briefly, 100 μL of p30 protein (5 μg/mL) were run in SDS-PAGE on a 12.5% polyacrylamide gel and electro-transferred to polyvinylidene fluoride (PVDF) membranes (Nylonmembran Immobilon-P, Millipore). After blocking with PBS-T/2% gelatin blocking buffer, the blotted membranes were incubated with sera and the same controls used for the ELISA diluted 1:100 in blocking buffer, followed by incubation with peroxidase conjugated AffiniPure rabbit anti-bovine IgG [H+L] (Jackson ImmunoResearch Laboratories (1:500) and developed with a substrate solution prepared with 4-chloro-1-naphthol (Sigma) and hydrogen peroxide.
2.2.4. IFAT
IFAT slides were prepared as described in previous work [21]. Briefly, T. gondii tachyzoites were grown in Vero cell (African Green Monkey kidney epithelial cells) cultures using Dulbecco’s Minimal Essential Medium (DMEM) supplemented with 100 UI mL−1 penicillin, 100 μg mL−1 streptomycin, 2 mM l-glutamine, and fetal calf serum (10% to initiate cultures and 2% for maintenance) in ventilated 75 cm2 cell culture flasks in a CO2 incubator at 37 °C. The day before harvesting of tachyzoites, fetal calf serum was withdrawn from the cell culture medium in order to reduce non-specific binding of antibodies. Tachyzoites were purified by filtration through Whatman CF11 cellulose columns [22] and fixed in 4% phosphate-buffered formaldehyde solution. After washing, the antigen suspension was diluted in PBS to a concentration of 2 × 106 tachyzoites mL−1 and used to coat IFAT slides. Serum samples were tested for specific anti-T. gondii antibodies at 1:50 dilution using anti-bovine IgG (whole molecule) FITC secondary antibody produced in rabbit (Sigma Aldrich, St. Louis, MO, USA) diluted at 1:200 in 0.01% Evans blue solution. Positive and negative sera from previous ELISA and WB analysis were included in each screening round.
2.3. Data Analysis
For statistical analysis purposes, an animal was considered seropositive for T. gondii if positive or suspicious results by the p30 ELISA were confirmed either by the p30 WB or IFAT and seropositive for N. caninum if positive by the p38 ELISA. Apparent animal-level and herd-level prevalence values were calculated with EpiTools Epidemiological Calculator using Wilson’s score for the 95% confidence intervals (CI) [23]. Variables evaluated as potential risk factors included age, herd size, human population density, Csa and Csb Köppen climate areas [24], farm location at NUTSII level, production system (intensive and extensive), production type (dairy and beef), and mean temperature and precipitation values between 1970 and 2000 (bio1–bio19). Climate data were obtained from the Worldclim 2.1 [25] dataset at a resolution of a 1000 m radius from the farm location and human population density (inhabitants/km2) was calculated at the municipality level, based on the population census 2011 and the Official Administrative Map of Portugal 2011. For ease of interpretation, the following continuous variables were categorized: age (<24 months and >24 months), herd size (≤50 animals and >50 animals to represent small-scale and large-scale farms), and population density (<50, 50–300, 300–1500, and >1500 inhabitants/km2, representing very low, low, semi-dense, and dense population, respectively [26]). Frequency distributions and statistical relationships were analyzed using the statistical software package R 4.2.0 [27]. Statistical significance of associations between serological results and explanatory variables was assessed by a binomial generalized linear mixed model fit by maximum likelihood (Laplace approximation) using the glmer function (lme4 package in R). Both univariate and multivariate analyses were performed. For the univariate analysis, each variable was entered individually into the model, while the multivariate model included all the significant factors from the univariate analysis. The variable “Farm ID” was set as a random factor in both analyses to control for possible variation due to particular characteristics of farms. Statistical significance was set at p < 0.05.
2.4. Spatial Analysis
Geographical distribution maps were constructed using Quantum Geographic Information System (QGIS) software 3.4. Spatial clustering of T. gondii and N. caninum seropositive farms was analyzed with the free SaTScan™ software version 9.7. using the Bernoulli probability model for high rates [28]. The following input data were considered: the number of animals with positive and negative results, and the latitude and longitude coordinates of each farm. The model was run with a circular spatial shape window and the default maximum spatial cluster size of 50% of the total population at risk.
3. Results
3.1. Descriptive Data
The 612 samples included in the survey were obtained in 35 farms located in 31 civil parishes, representing 26 municipalities in four of the five NUTS II regions of continental Portugal: Norte (n = 259), Alentejo (n = 185), Centro (n = 148), and Lisboa (n = 20) (Table 1). The 35 herds in the study displayed a median of 102 animals (IQR = 51–211.8), with a minimum of 29 and a maximum of 540 animals. The median age of cattle was 58 months (IQR = 38–87), with a minimum of 9 and a maximum of 240 months. There were 5 sera from male and 607 from female cattle.
3.2. N. caninum Seroprevalence Results
Antibodies to N. caninum were detected in 105 (17.2%; 95% CI 14.4–20.4) of the sera tested using the p38 ELISA. At farm level, at least one positive animal was detected in 24 farms (68.6%; 95% CI 52.0–81.5%). Positive farms were located in 21 civil parishes (70%) in 17 municipalities (70.8%) included in the study. Animal and herd-level N. caninum prevalence according to the categorical variables age, sex, herd size, production system, production type, Köppen climate area, NUTS2 region, and population density considered in the risk factor analysis is shown in Table 1.
3.3. T. gondii Seroprevalence Results
Using the p30 ELISA, 31 (5.1%) of the 612 sera were classified as positive at the 95th percentile and further 31 sera were classified as suspicious at the 90th percentile cut-off. Confirmation of p30 ELISA positive and suspicious results either by the p30 WB or IFAT confirmed T. gondii seropositivity in 56 animals, representing a seroprevalence of 9.2% (95% CI 7.1–11.7%) (Table 1). Concerning the panel of 31 negative sera, 2 showed a positive reaction by the p30 WB, while none was reactive by IFAT. At farm level, at least one positive animal was detected by the p30 ELISA in 14 farms (40%) when using the 95th percentile as the cut-off for seropositivity and at least one suspicious animal was identified in further 5 farms when using the 90th percentile. A total of 18 farms (51.4%; 95% CI 35.6–67.0%) were found to be positive when confirming positive and suspicious results by the p30WB or IFAT (Table 1). Positive farms were distributed in 17 civil parishes (56.7%) in 14 municipalities (58.3%) included in the study. Distribution of animal and herd-level T. gondii prevalence according to the categorical variables age, sex, herd size, human population density, Köppen climate area, NUTS2 region, production system and production type considered in the risk factor analysis is shown in Table 1.
3.4. Sera Positive for Both, N. caninum and T. gondii
Of the 105 sera positive for N. caninum, 11 were also positive for T. gondii. Sera positive for both parasites were from 5 farms. A total of 7 of the T. gondii positive sera were confirmed both by p30 WB and IFAT in the 5 farms, 2 sera were confirmed by IFAT only, and other 2 sera by p30 WB only in 3 of the farms.
3.5. Univariate Analysis
3.5.1. Risk Factors for N. caninum Seroprevalence
Based on the univariate analysis, human population density, Köppen climate area, NUTS2 region, production system, and bioclimatic variables bio13—precipitation of wettest month, bio14—precipitation of driest month, bio17—precipitation of driest quarter, and bio18—precipitation of warmest quarter were significantly associated with N. caninum seroprevalence (Table 2). Hence, cattle raised in semi-densely (300–500 inhabitants/km2) and densely (>1500 inhabitants/km2) populated municipalities were 4.6 (OR = 4.646; 95%CI 1.547–13.949) and 13.64 times (OR = 13.64; 95% CI 2.086–89.176) more likely to be N. caninum positive than cattle in areas with a low or very low population density. The odds of infection were higher for cattle raised in Csb climate areas (OR = 4.352; 95% CI 1.411–13.421) compared to Csa areas and higher in the NUTS2 region Norte (OR = 7.11; 95% CI 1.836–27.537) compared to the regions Centro, Lisboa, and Alentejo. In addition, animals raised in intensive production systems were more likely to be positive (OR = 6.396; 95% CI 1.831–22.334), when compared to extensively raised cattle. The odds ratios determined for bioclimatic precipitation variables bio13 (OR = 1.024; 95% CI 1.007–1.041), bio14 (OR = 1.154; 95% CI 1.037–1.283), bio17 (OR = 1.032; CI 1.001–1.057), and bio18 (OR = 1.028; 1.001–1.047) showed a similar positive effect on infection prevalence. The variables age, herd size, and production type (Table 1) were not significantly associated to an increased risk of infection. No statistically significant relationship was found between N. caninum and T. gondii seropositivity.
3.5.2. Risk Factors for T. gondii Seroprevalence
Based on the univariate analysis, herd size, human population density, climate area, and bioclimatic variables bio13—precipitation of wettest month, bio 14—precipitation of driest month, bio17—precipitation of driest quarter, and bio18—precipitation of warmest quarter were identified as significant predictors of T. gondii seroprevalence (Table 3). Thus, cattle in large herds (≥50 animals) had a lower risk of T. gondii infection (OR = 0.126) compared to animals in small herds, while human population density above 1500 inhabitants/km2 increased the odds of infection by 15.786 times (OR = 15.786; 95% CI 1.682–148.139). The probability of being seropositive was higher for cattle in Csb climate areas compared to Csa areas (OR = 4.521; 95% CI 1.269–16.109). The odds ratios determined for bioclimatic precipitation variables bio13 (OR = 1.026; 95% CI 1.007–1.045), bio14 (OR = 1.149; 95% CI 1.022–1.292), bio17 (OR = 1.031; 95% CI 1.004–1.058), and bio18 (OR = 1.025; 95% CI 1.004–1.047) showed a similar positive effect on infection prevalence. The variables age, NUTS2 region, production system, and production type (Table 1) were not significantly associated with an increased risk of infection.
3.6. Multivariable Risk Factor Analysis
None of the significant factors in the univariate analysis was associated with a higher risk of infection with T. gondii or N. caninum in the final multivariate model.
3.7. Spatial Analysis Results
- N. caninum
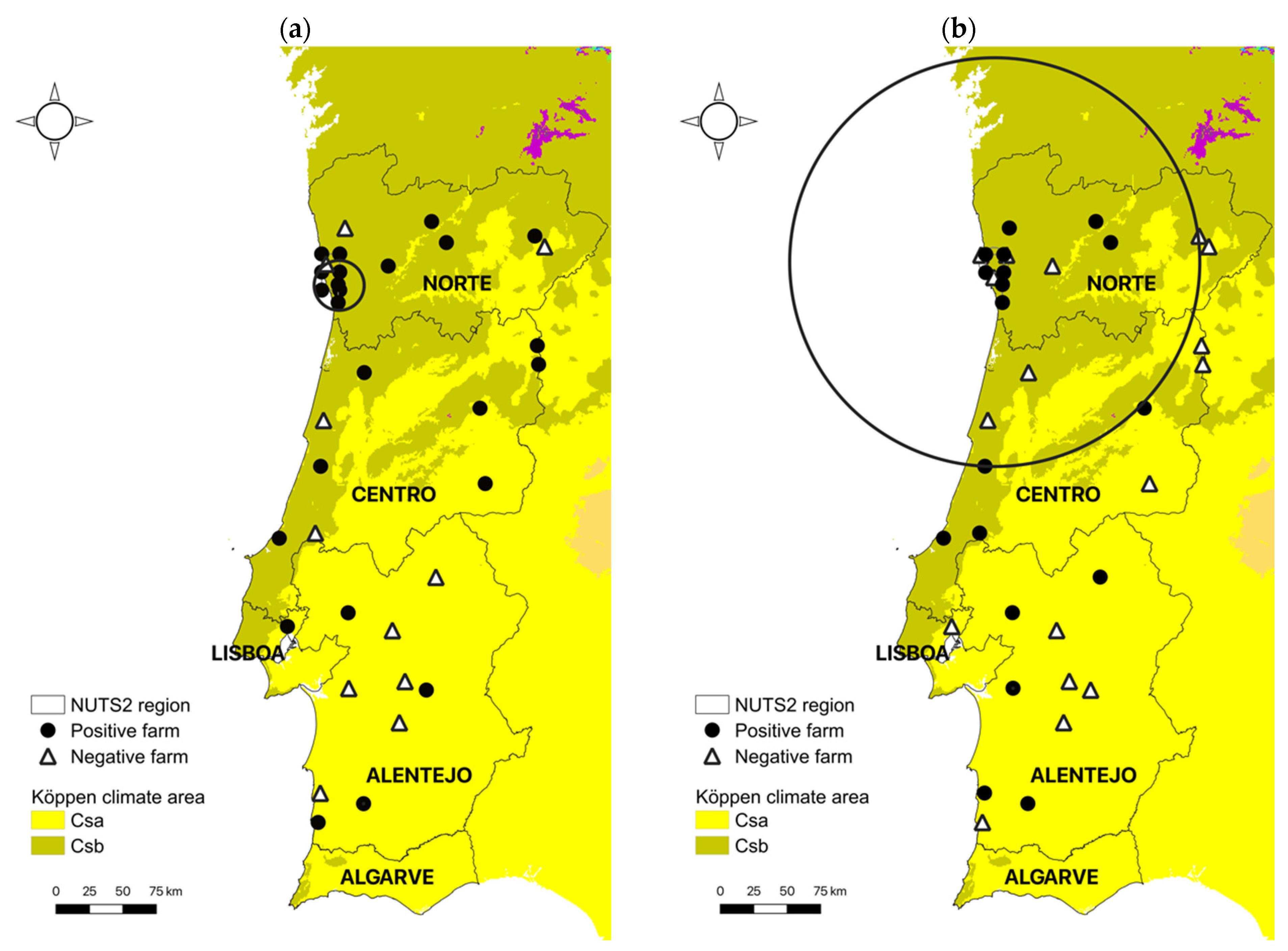
Spatial cluster analysis identified one significant cluster (p < 0.0001) of high occurrence of N. caninum infection (Figure 1a). The cluster included a total of 54 positive animals out of 128 in 6 of 7 farms and the resulting cluster radius was 18.7 km. The proportion of positive cases within the cluster area was 42.2% and the relative risk of an individual animal being infected with N. caninum compared to animals outside the cluster was 4.0.
- T. gondii
Spatial cluster analysis identified one significant cluster (p < 0.0001) of high occurrence of T. gondii infection (Figure 1b). The cluster included a total of 43 positive animals out of 261 in 10 out of 16 farms and the resulting cluster radius was 152.9 km. The proportion of positive cases within the cluster area was 16.5% and the relative risk of an individual animal being infected with T. gondii compared to animals outside the cluster was 4.5.
4. Discussion
The results of this study show a relatively low but widespread prevalence of T. gondii infection in cattle in Portugal. The overall seroprevalence of 8.0% and, in particular, the seroprevalence of 12.0% determined in the NUTS2 region Norte are consistent with previous reports from Portugal (7.5%) [30] and the neighboring Spanish region of Galicia (7.3%) [31]. However, because two of the sera regarded as negative in the ELISA tested positive in one of the tests used for confirmation, it cannot be excluded that the overall T. gondii prevalence in Portugal is higher than estimated here. Though similar low proportions of seropositivity (3.2–9.7%) were reported in several surveys in cattle across Europe over the past two decades [32,33,34], exposure to T. gondii seems to vary considerably, and other studies point to proportions of seropositive cattle up to 83.3% [35,36,37,38,39]. Regarding seroprevalence results for N. caninum, considering only studies in which larger numbers of bovine herds were involved, a higher N. caninum seroprevalence in southern Europe compared to northern Europe seems to be apparent (Portugal 28.0% [40], Spain 22.5–22.6% [41], Switzerland 11.5% [42], Belgium 10.2% [43], the Netherlands 9.9% [44], Germany 4.0%, 4.1% or 6.8% [44,45,46], Poland 19.5% [47], Sweden 1.3% or 2.8% [44,48]).
Considering life-cycle characteristics and conditions that might influence the viability of oocysts in soil, differences in the prevalence of both parasites may result from a variety of factors assessed in the present survey, including climate, geographical region, management conditions, herd size, age of animals, human density, or a combination thereof [5,49]. Further, prevalence may also vary according to the type of sample and diagnostic test used.
Regarding climatic factors, weather conditions in continental Portugal are influenced by the latitude and proximity to the Atlantic Ocean and some climatic variables, such as precipitation and temperature, present strong north–south and west–east gradients. According to Köppen’s classification, the climate in Portugal can be grossly divided in two distinct temperate regions: one characterized by rainy winters and dry, hot summers (Csa climate area) in the southern part of the country, and the other by rainy winters and dry, mild summers (Csb climate area) in the north (Figure 1) [29]. In the present study, infection with both parasites was more likely in Csb climate areas, especially in the northwest of Portugal (Table 1, Figure 1). The prevalence of T. gondii in soil was shown to be higher in wet or moist seasons characterized by mild temperatures [50] and both sporulation and viability of sporulated oocysts are compromised by extreme heat (>45 °C), and desiccation [51,52,53]. Little is known about the survival of N. caninum oocysts in soil, but due to its phylogenetic resemblance with T. gondii, a similar resistance to environmental conditions is presumed. Thus, weather conditions in Csb areas in the north of Portugal are expected to be more favorable for oocyst survival across seasons, as opposed to the extreme hot and dry conditions frequently observed during the summer in the Centro-Alentejo area. Further supporting this relationship, when looking more specifically at bioclimatic variables characterizing Csa and Csb climate areas, the ones positively associated with an increased risk of N. caninum and T. gondii infection were identical and related to precipitation during extreme environmental conditions (wettest and driest month and driest and warmest quarter). In a serological survey on T. gondii prevalence in the human Portuguese population comparing three cross-sectional studies spanning three decades (1979–2013), a higher seroprevalence was also observed in the NUTS2 region Norte in the first two decades, although this trend seemed to be inverted in the last decade, with an unexpected raise in the south [54]. The reasons for these contradicting findings, e.g., a change in food habits, with an increase in the consumption or preference for rare beef, still need to be clarified. As for N. caninum, the higher infection rates found in the NUTS2 region Norte in the present study (Table 1) reflect previous data in Portugal showing a seroprevalence of 28% in areas of intensive dairy production in the regions Norte and Centro of Portugal [40].
In addition to climate, regional differences in cattle husbandry and management practices are other factors that may contribute to the spatial clustering of T. gondii and N. caninum infection in the north of Portugal. Thus, in Portugal, dairy production is concentrated in the northwest mainland area, where cows are reared intensively, mostly under permanent confinement conditions. In intensively managed cattle, stocking density and increased potential for contamination of feeding areas, water, fodder, stored silage, and feed supplements by dog feces has been linked to an increased risk of postnatal N. caninum infection [49] and could explain the higher prevalence of this parasite in intensive production systems in the present study. On the other hand, the higher internal replacement rates in dairy cattle farms in Portugal compared to beef cattle could also explain the higher prevalence of N. caninum through vertical transmission, though the differences for production type (dairy vs. beef) did not reach statistical significance in the GLMM model.
Notably, in a study conducted in cats in France, seroprevalence was increased in areas with high farm densities and during years with cool and moist winters, also showing the effect of climate and farming conditions on the prevalence of T. gondii in the definitive host [55]. In the present study, the seroprevalence of both parasites was higher in small-scaled herds, but herd size was found to be a risk factor only for T. gondii. Smaller herds are associated to more traditional ways of farming and may favor contamination of feed and water with T. gondii oocysts, for instance, through a higher density of cats and lack of biosecurity measures. In fact, studies carried out in several livestock species, including cattle, show that the smaller the herd or flock, the higher the chance for seropositivity to T. gondii [5,32,33,37,39]. In the case of N. caninum, both risk and protective effects were reported for herd size [49,56].
A possible relationship with indirect risk factors such as human population density, which may play a role in T. gondii and N. caninum transmission as a result of increased cat and dog populations [53,57], was also assessed in the present survey. Results from this analysis show that the risk of infection was positively associated with increasing population density for both parasites as observed elsewhere [57]. This notwithstanding, T. gondii and N. caninum transmission dynamics may be influenced by other factors such as heterogeneity in land use and variations in intermediate host and definitive host populations along the urban–rural–wild gradient [58], which need to be addressed in more detail.
Though seroprevalence of T. gondii and N. caninum is expected to increase with age due to an assumed lifelong persistence of tissue cysts, as noted by several authors [30,33,36,49,59,60], others have demonstrated the opposite [61,62] or that age-prevalence may be influenced by other factors, for instance by the presence of cats in the case of T. gondii, with higher seroprevalences in younger animals in farms with cats and in older cows in farms without cats [32]. Here, the lower seroprevalence in animals aged <24 months was not statistically significant. Possible explanations could be the absence of a lifelong antibody response to both parasites in cattle, age-related differences in immune response, or the decrease over time of antibody titers to levels below the detection limit of the serological technique used. Adding to this, cut-off values reported in literature for the serological screening of cattle vary substantially between studies [34,36,49,63,64]. Thus, the presence of low antibody titers in adult cattle could go unnoticed when using more conservative cut-off thresholds in serological surveys. Finally, the serological test itself may have a significant effect on prevalence rates both between and within age groups. Though several tests for the serological diagnosis of T. gondii and N. caninum were already developed for cattle, few attempts were made to compare their performance in terms of sensitivity and specificity and only a minority of surveys included a second confirmatory test. In the absence of a gold standard test, serial testing approaches may be more advantageous for surveys of diseases with an expected low prevalence, by improving specificity with moderate loss in sensitivity and increasing the positive predictive value of results. Therefore, in regard to T. gondii, the strategy in this study was to enhance sensitivity by including a cut-off at the 90th percentile for the p30 ELISA in order to identify suspicious sera, followed by confirmation by two different serological tests to warrant specificity of results. The observation that only a minority of sera were double positive for closely related parasites, T. gondii and N. caninum, provides confidence that the positive reactions in the T. gondii test are true positives and have not been caused by cross-reactions. Interestingly, double positives clustered in a limited number of herds, which shows that specific conditions, probably due to particular epidemiological situations (e.g., presence of dogs and cats) may have led to co-infection.
5. Conclusions
Data obtained in the present study may be valuable as input for quantitative source attribution models to assess the likely contribution of beef as a source of T. gondii infection in humans. The prevalence determined for N. caninum confirms the need to include neosporosis in the differential diagnosis of abortive or reproductive disorders in cattle in Portugal. The risk factors and spatial clustering of infection seen in the present survey for both parasites highlight the importance of biosecurity measures in cattle farms to avoid contamination of feed and water with dog and cat feces, in particular in areas where management and husbandry practices may favor transmission and climate conditions are suited to the survival of oocysts in the environment.
Author Contributions
Conceptualization, G.S. and H.W.; methodology A.B. and H.W.; validation, G.S., T.N. and H.W.; investigation, G.S., T.N., A.B. and H.W.; resources, G.S. and H.W.; data curation, G.S., T.N. and H.W.; writing—original draft preparation, H.W.; writing—review and editing, G.S. and T.N.; supervision, G.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to the retrospective character of the study, based on sera selected from a serum collection obtained during a previous survey [14].
Informed Consent Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analyzed in this study. Data on cattle population in Portugal can be found at https://dgav.pt, accessed on 2 March 2022. Mean temperature and precipitation values between 1970 and 2000 (bio1–bio19) were obtained from the Worldclim 2.1 dataset available at https://www.worldclim.org/data/worldclim21.html, accessed on 4 October 2021. Human population density at the municipality level (inhabitants/km2) was calculated based on census data available at https://censos.ine.pt/xportal/xmain?xpid=CENSOS&xpgid=censos_ficheirosintese, accessed on 2 March 2022, and municipality areas provided in the Official Administrative Map of Portugal CAOP 2011 in https://www.dgterritorio.gov.pt/cartografia/cartografia-tematica/caop, accessed on 2 March 2022. The map in Figure 1 was constructed in QGIS 3.4 Open Source Geographic Information System using freely available layers: administrative boundaries for NUTS2 regions were obtained at https://ec.europa.eu/eurostat/web/gisco/geodata/reference-data/administrative-units-statistical-units, accessed on 2 March 2022, the Köppen-Geiger classification map for climate areas (Beck_KG_V1_present_0p0083.tif) is freely available for download at http://www.gloh2o.org/koppen/, accessed on 24 September 2021.
Acknowledgments
The authors thank Lucinda Marques and Maria do Carmo Ramos (INIAV/Parasitology Laboratory) for valuable laboratory assistance. This work resulted from a collaboration within the framework of Euro-FBP, the European Network for Foodborne Parasites, COST Action FA1408.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dubey, J. Review of Neospora caninum and neosporosis in animals. Korean J. Parasitol. 2003, 41, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Thurmond, M.C.; Hietala, S.K. Effect of congenitally acquired Neospora caninum infection on risk of abortion and subsequent abortions in dairy cattle. Am. J. Vet. Res. 1997, 58, 1381–1385. [Google Scholar] [PubMed]
- Barling, K.S.; McNeill, J.W.; Thompson, J.A.; Paschal, J.C.; McCollum, F.T.; Craig, T.M.; Adams, L.G. Association of serologic status for Neospora caninum with postweaning weight gain and carcass measurements in beef calves. J. Am. Vet. Med. Assoc. 2000, 217, 1356–1360. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.; Risco, C.; Donovan, A. Association between exposure to Neospora caninum and milk production in dairy cows. J. Am. Vet. Med. Assoc. 2001, 219, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, S.; Basso, W.; Benavides Silván, J.; Ortega-Mora, L.M.; Maksimov, P.; Gethmann, J.; Conraths, F.J.; Schares, G. Toxoplasma gondii infection and toxoplasmosis in farm animals: Risk factors and economic impact. Food Waterborne Parasitol. 2019, 15, e00037. [Google Scholar] [CrossRef]
- Delair, E.; Monnet, D.; Grabar, S.; Dupouy-Camet, J.; Yéra, H.; Brézin, A. Acquired and Congenital Ocular Toxoplasmosis in a French Population. Invest. Ophthalmol. Vis. Sci. 2006, 47, 293. [Google Scholar]
- Petersen, E.; Kijlstra, A.; Stanford, M. Epidemiology of ocular toxoplasmosis. Ocul. Immunol. Inflamm. 2012, 20, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Fekadu, A.; Shibre, T.; Cleare, A.J. Toxoplasmosis as a cause for behaviour disorders—Overview of evidence and mechanisms. Folia Parasitol. (Praha) 2013, 57, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.J.C.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of toxoplasma infection in pregnant women: European multicentre case-control study. BMJ 2000, 321, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Kotula, A.W.; Dubey, J.P.; Sharar, A.K.; Andrews, C.D.; Shen, S.K.; Lindsay, D.S. Effect of freezing on infectivity of Toxoplasma gondii tissue cysts in pork. J. Food Prot. 1991, 54, 687–690. [Google Scholar] [CrossRef]
- El-Nawawi, F.A.; Tawfik, M.A.; Shaapan, R.M. Methods for inactivation of Toxoplasma gondii cysts in meat and tissues of experimentally infected sheep. Foodborne Pathog. Dis. 2008, 5, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Opsteegh, M.; Prickaerts, S.; Frankena, K.; Evers, E.G. A quantitative microbial risk assessment for meatborne Toxoplasma gondii infection in The Netherlands. Int. J. Food Microbiol. 2011, 150, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H.; Yang, Y.R. Public Health Significance of Toxoplasma gondii Infections in Cattle: 2009–2020. J. Parasitol. 2020, 106, 772–788. [Google Scholar] [CrossRef] [PubMed]
- Waap, H.; Nunes, T.; Cortes, H.; Leitão, A.; Vaz, Y. Prevalence and geographic distribution of Besnoitia besnoiti infection in cattle herds in Portugal. Parasitol. Res. 2014, 113, 3703–3711. [Google Scholar] [CrossRef]
- Dohoo, I.; Martin, W.; Stryhn, H. Veterinary Epidemiological Research, 1st ed.; AVC Inc.: Charlottetown, PE, Canada, 2003; pp. 27–52. [Google Scholar]
- Schares, G.; Rauser, M.; Söndgen, P.; Rehberg, P.; Bärwald, A.; Dubey, J.P.; Edelhofer, R.; Conraths, F.J. Use of purified tachyzoite surface antigen p38 in an ELISA to diagnose bovine neosporosis. Int. J. Parasitol. 2000, 30, 1123–1130. [Google Scholar] [CrossRef]
- Pardini, L.; Maksimov, P.; Herrmann, D.C.; Bacigalupe, D.; Rambeaud, M.; Machuca, M.; Moré, G.; Basso, W.; Schares, G.; Venturini, M.C. Evaluation of an in-house TgSAG1 (P30) IgG ELISA for diagnosis of naturally acquired Toxoplasma gondii infection in pigs. Vet. Parasitol. 2012, 189, 204–210. [Google Scholar] [CrossRef]
- Seltmann, A.; Schares, G.; Aschenborn, O.H.K.; Heinrich, S.K.; Thalwitzer, S.; Wachter, B.; Czirják, G.Á. Species-specific differences in Toxoplasma gondii, Neospora caninum and Besnoitia besnoiti seroprevalence in Namibian wildlife. Parasit. Vectors 2020, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Lücht, M.; Stagegaard, J.; Conraths, F.J.; Schares, G. Toxoplasma gondii in small exotic felids from zoos in Europe and the Middle East: Serological prevalence and risk factors. Parasit. Vectors 2019, 12, 449. [Google Scholar] [CrossRef]
- Opsteegh, M.; Spano, F.; Aubert, D.; Balea, A.; Burrells, A.; Cherchi, S.; Cornelissen, J.B.W.J.; Dam-Deisz, C.; Guitian, J.; Györke, A.; et al. The relationship between the presence of antibodies and direct detection of Toxoplasma gondii in slaughtered calves and cattle in four European countries. Int. J. Parasitol. 2019, 49, 515–522. [Google Scholar] [CrossRef]
- Waap, H.; Volkart de Oliveira, U.; Nunes, T.; Gomes, J.; Gomes, T.; Bärwald, A.; Dias Munhoz, A.; Schares, G. Serological survey of Neospora spp. and Besnoitia spp. in horses in Portugal. Vet. Parasitol. Reg. Stud. Rep. 2020, 20, 100391. [Google Scholar] [CrossRef]
- Dempster, R.P. Toxoplasma gondii: Purification of zoites from peritoneal exudates by eight methods. Exp. Parasitol. 1984, 57, 195–207. [Google Scholar] [CrossRef]
- Sergeant, E.S.G. Epitools Epidemiological Calculators. Ausvet. 2018. Available online: http://epitools.ausvet.com.au (accessed on 10 October 2021).
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen–Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- European Union; FAO; UN-Habitat; OECD; The World Bank. Applying the Degree of Urbanisation: A Methodological Manual to Define Cities, Towns and Rural Areas for International Comparisons. OECD Regional Development Studies; OECD Publishing: Paris, France; European Union: Brussels, Belgium, 2021. [Google Scholar] [CrossRef]
- Bunn, A.; Korpela, M. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. 2013. Available online: https://cran.microsoft.com/snapshot/2014-09-08/web/packages/dplR/vignettes/xdate-dplR.pdf (accessed on 10 October 2021).
- Kulldorff, M. A spatial scan statistic. Commun. Stat. Theory Methods 1997, 26, 1481–1496. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger 584 climate classification maps at 1-km resolution. Sci. Data. 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.P.; Dubey, J.P.; Neto, F.; Rodrigues, A.; Martins, T.; Rodrigues, M.; Cardoso, L. Seroprevalence of Toxoplasma gondii infection in cattle, sheep, goats and pigs from the North of Portugal for human consumption. Vet. Parasitol. 2013, 193, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Panadero, R.; Painceira, A.; López, C.; Vázquez, L.; Paz, A.; Díaz, P.; Dacal, V.; Cienfuegos, S.; Fernández, G.; Lago, N.; et al. Seroprevalence of Toxoplasma gondii and Neospora caninum in wild and domestic ruminants sharing pastures in Galicia (Northwest Spain). Res. Vet. Sci. 2010, 88, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Gilot-Fromont, E.; Aubert, D.; Belkilani, S.; Hermitte, P.; Gibout, O.; Geers, R.; Villena, I. Landscape, herd management and within-herd seroprevalence of Toxoplasma gondii in beef cattle herds from Champagne-Ardenne, France. Vet. Parasitol. 2009, 161, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gąsior, L.; Drapała, D.; Dominiak-Górski, B.; Kur, J. Epidemiological study of Toxoplasma gondii infection among cattle in Northern Poland. Ann. Agric. Environ. Med. 2013, 20, 653–656. [Google Scholar] [PubMed]
- Jokelainen, P.; Tagel, M.; Mõtus, K.; Viltrop, A.; Lassen, B. Toxoplasma gondii seroprevalence in dairy and beef cattle: Large-scale epidemiological study in Estonia. Vet. Parasitol. 2017, 236, 137–143. [Google Scholar] [CrossRef]
- Berger-Schoch, A.E.; Bernet, D.; Doherr, M.G.; Gottstein, B.; Frey, C.F. Toxoplasma gondii in Switzerland: A serosurvey based on meat juice analysis of slaughtered pigs, wild boar, sheep and cattle. Zoonoses Public Health 2011, 58, 472–478. [Google Scholar] [CrossRef]
- Blaga, R.; Aubert, D.; Thébault, A.; Perret, C.; Geers, R.; Thomas, M.; Alliot, A.; Djokic, V.; Ortis, N.; Halos, L.; et al. Toxoplasma gondii dans la viande bovine consommée en France: Variation régionale de la séroprévalence et isolement de parasites. Parasite Paris Fr. 2019, 26, 77. [Google Scholar] [CrossRef] [PubMed]
- García-Bocanegra, I.; Cabezón, O.; Hernández, E.; Martínez-Cruz, M.S.; Martínez-Moreno, Á.; Martínez-Moreno, J. Toxoplasma gondii in ruminant species (cattle, sheep, and goats) from southern Spain. J. Parasitol. 2013, 99, 438–440. [Google Scholar] [CrossRef]
- Klun, I.; Djurković-Djaković, O.; Katić-Radivojević, S.; Nikolić, A. Cross-sectional survey on Toxoplasma gondii infection in cattle, sheep and pigs in Serbia: Seroprevalence and risk factors. Vet. Parasitol. 2006, 135, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Sroka, J. Seroepidemiology of toxoplasmosis in the Lublin region. Ann. Agric. Environ. Med. 2001, 8, 25–31. [Google Scholar]
- Canada, N.; Carvalheira, J.; Meireles, C.S.; Correia da Costa, J.M.; Rocha, A. Prevalence of Neospora caninum infection in dairy cows and its consequences for reproductive management. Theriogenology 2004, 62, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Eiras, C.; Arnaiz, I.; Alvarez-García, G.; Ortega-Mora, L.M.; Sanjuánl, M.L.; Yus, E.; Diéguez, F.J. Neospora caninum seroprevalence in dairy and beef cattle from the northwest region of Spain, Galicia. Prev. Vet. Med. 2011, 98, 128–132. [Google Scholar] [CrossRef]
- Gottstein, B.; Hentrich, B.; Wyss, R.; Thür, B.; Busato, A.; Stärk, K.D.; Müller, N. Molecular and immunodiagnostic investigations on bovine neosporosis in Switzerland. Int. J. Parasitol. 1998, 28, 679–691. [Google Scholar] [CrossRef]
- Vangeel, I.; Méroc, E.; Roelandt, S.; Welby, S.; Riocreux, F.; Stede, Y.V.D.; Driessche, E.V.; Schoubroeck, L.V.; Czaplicki, G.; Quinet, C.; et al. Seroprevalence of Neospora caninum, paratuberculosis and Q fever in cattle in Belgium in 2009–2010. Vet. Rec. 2012, 171, 477. [Google Scholar] [CrossRef]
- Bartels, C.J.M.; Arnaiz-Seco, J.I.; Ruiz-Santa-Quitera, A.; Björkman, C.; Frössling, J.; von Blumröder, D.; Conraths, F.J.; Schares, G.; van Maanen, C.; Wouda, W.; et al. Supranational comparison of Neospora caninum seroprevalences in cattle in Germany, The Netherlands, Spain and Sweden. Vet. Parasitol. 2006, 137, 17–27. [Google Scholar] [CrossRef] [PubMed]
- von Blumröder, D.; Stambusch, R.; Labohm, R.; Klawonn, W.; Dräger, K.; Fasen, W.; Conraths, F.J.; Schares, G. Potenzielle Risikofaktoren für den serologischen Nachweis von Neospora-caninum-Infektionen in Rinderherden in Rheinland-Pfalz. Tierärztl. Prax. Ausg. G Großtiere Nutztiere 2006, 34, 141–147. [Google Scholar] [CrossRef]
- Weber, A.; Zetzmann, K.; Ewringmann, T. Vorkommen von Antikörpern gegen Neospora caninum bei Kühen in nordbayerischen Beständen mit Abortproblemen. Tierärztliche Umsch. 2000, 55, 27–29. [Google Scholar]
- Kowalczyk, S.J.; Czopowicz, M.; Weber, C.N.; Müller, E.; Witkowski, L.; Kaba, J. Herd-level seroprevalence of Neospora caninum infection in dairy cattle in central and northeastern Poland. Acta Parasitol. 2016, 61, 63–65. [Google Scholar] [CrossRef]
- Loobuyck, M.; Frössling, J.; Lindberg, A.; Björkman, C. Seroprevalence and spatial distribution of Neospora caninum in a population of beef cattle. Prev. Vet. Med. 2009, 92, 116–122. [Google Scholar] [CrossRef]
- Dubey, J.P.; Schares, G.; Ortega-Mora, L.M. Epidemiology and Control of Neosporosis and Neospora caninum. Clin. Microbiol. Rev. 2007, 20, 323–367. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Feng, H.L.; Nie, H.; Tu, P.; Zhang, Q.L.; Hu, M.; Zhou, Y.Q.; Zhao, J.L. Survey on the contamination of Toxoplasma gondii oocysts in the soil of public parks of Wuhan, China. Vet. Parasitol. 2012, 184, 141–146. [Google Scholar] [CrossRef]
- Dubey, J.P. Toxoplasma gondii Oocyst Survival under Defined Temperatures. J. Parasitol. 1998, 84, 862–865. [Google Scholar] [CrossRef] [PubMed]
- Lélu, M.; Villena, I.; Dardé, M.-L.; Aubert, D.; Geers, R.; Dupuis, E.; Marnef, F.; Poulle, M.-L.; Gotteland, C.; Dumètre, A.; et al. Quantitative estimation of the viability of Toxoplasma gondii oocysts in soil. Appl. Environ. Microbiol. 2012, 78, 5127–5132. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, K.; Bahia-Oliveira, L.; Dixon, B.; Dumètre, A.; de Wit, L.A.; VanWormer, E.; Villena, I. Environmental transmission of Toxoplasma gondii: Oocysts in water, soil and food. Food Waterborne Parasitol. 2019, 15, e00049. [Google Scholar] [CrossRef] [PubMed]
- Gargaté, M.J.; Ferreira, I.; Vilares, A.; Martins, S.; Cardoso, C.; Silva, S.; Nunes, B.; Gomes, J.P. Toxoplasma gondii seroprevalence in the Portuguese population: Comparison of three cross-sectional studies spanning three decades. BMJ Open 2016, 6, e011648. [Google Scholar] [CrossRef]
- Afonso, E.; Germain, E.; Poulle, M.L.; Ruette, S.; Devillard, S.; Say, L.; Villena, I.; Aubert, D.; Gilot-Fromont, E. Environmental determinants of spatial and temporal variations in the transmission of Toxoplasma gondii in its definitive hosts. Int. J. Parasitol Parasites Wildl. 2013, 2, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Hemphill, A.; Calero-Bernal, R.; Schares, G.; Hemphill, A.; Calero-Bernal, R.; Schares, G. Neosporosis in Animals; CRC Press: Boca Ratan, FL, USA, 2017. [Google Scholar] [CrossRef]
- Schares, G.; Bärwald, A.; Staubach, C.; Ziller, M.; Klöss, D.; Schröder, R.; Labohm, R.; Dräger, K.; Fasen, W.; Hess, R.G.; et al. Potential risk factors for bovine Neospora caninum infection in Germany are not under the control of the farmers. Parasitology 2004, 129, 301–309. [Google Scholar] [CrossRef]
- Gilot-Fromont, E.; Lélu, M.; Dardé, M.L.; Richomme, C.; Aubert, D.; Afonso, E.; Mercier, A.; Gotteland, C.; Villena, I. Toxoplasmosis—Recent Advances; IntechOpen: London, UK, 2012; Chapter 1; pp. 3–36. [Google Scholar] [CrossRef]
- Ahmad, N.; Qayyum, M. Seroprevalence and risk factors for toxoplasmosis in large ruminants in northern Punjab, Pakistan. J. Infect. Dev. Ctries. 2014, 8, 1022–1028. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, D.; Yang, S.-C.; Zhu, J.-H.; Li, J.-M.; Shi, K.; Du, R.; Zhao, Q. Prevalence and Risk Factors of Brucellosis, Toxoplasmosis, and Neosporosis Among Yanbian Yellow Cattle in Jilin Province, China. Vector Borne Zoonotic Dis. 2019, 19, 217–221. [Google Scholar] [CrossRef]
- Nematollah, A.; Moghddam, G.H. Survey on Seroprevalence of Anti-Toxoplasma gondii antibodies in cattle in Tabriz (Iran) by IFAT. Am. J. Anim. Vet. Sci. 2008, 3, 40–42. [Google Scholar] [CrossRef]
- Tialla, D.; Dahourou, L.D.; Gbati, O.B. Seroprevalence and factors associated with bovine and swine toxoplasmosis in Bobo-Dioulasso, Burkina Faso. Vet. World 2019, 12, 1519–1523. [Google Scholar] [CrossRef] [PubMed]
- Chandrawathani, P.; Nurulaini, R.; Zanin, C.M.; Premaalatha, B.; Adnan, M.; Jamnah, O.; Khor, S.K.; Khadijah, S.; Lai, S.Z.; Shaik, M.; et al. Seroprevalence of Toxoplasma gondii antibodies in pigs, goats, cattle, dogs and cats in peninsular Malaysia. Trop. Biomed. 2008, 25, 257–258. [Google Scholar] [PubMed]
- Fajardo, H.V.; D’ávila, S.; Bastos, R.R.; Cyrino, C.D.; de Lima Detoni, M.; Garcia, J.L.; das Neves, L.B.; Nicolau, J.L.; Amendoeira, M.R.R. Seroprevalence and risk factors of toxoplasmosis in cattle from extensive and semi-intensive rearing systems at Zona da Mata, Minas Gerais state, Southern Brazil. Parasites Vectors 2013, 6, 191. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Locations of Neospora caninum (a) and Toxoplasma gondii (b) positive and negative farms in the different NUTS2 regions in continental Portugal. Significant clusters of infection detected by spatial cluster analysis represented by circles (small circle with arrow for N. caninum and large circle for T. gondii). Csa and Csb Köppen climate areas according to Beck et al. 2018 [29]. © EuroGeographics for the administrative boundaries.
Figure 1.
Locations of Neospora caninum (a) and Toxoplasma gondii (b) positive and negative farms in the different NUTS2 regions in continental Portugal. Significant clusters of infection detected by spatial cluster analysis represented by circles (small circle with arrow for N. caninum and large circle for T. gondii). Csa and Csb Köppen climate areas according to Beck et al. 2018 [29]. © EuroGeographics for the administrative boundaries.


{kind=link}
Table 1.
Animal and herd seroprevalence (%) of Toxoplasma gondii and Neospora caninum in cattle in Portugal according to age, sex, herd size, human population density, Köppen climate area, NUTSII regions, production system, and production type. (CI) Confidence Interval for prevalence.
Table 1.
Animal and herd seroprevalence (%) of Toxoplasma gondii and Neospora caninum in cattle in Portugal according to age, sex, herd size, human population density, Köppen climate area, NUTSII regions, production system, and production type. (CI) Confidence Interval for prevalence.
| Animal Seroprevalence | Herd Seroprevalence | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T. gondii | N. caninum | T. gondii | N. caninum | ||||||||
| Variables | N | Positive | % (CI) | Positive | % (CI) | N | Positive | % (CI) | Positive | % (CI) | |
| Age | ≤24 | 52 | 4 | 7.7 (3.0–18.2) | 3 | 5.8 (2.0–15.6) | |||||
| >24 | 560 | 52 | 9.3 (7.2–11.0) | 102 | 18.2 (15.2–21.6) | ||||||
| Sex | Female | 607 | 55 | 9.1 (7.0–11.6) | 105 | 17.3 (14.5–20.5) | |||||
| Male | 5 | 1 | 20.0 (3.6–62.5) | 0 | |||||||
| Herd size | ≤50 animals | 144 | 34 | 23.6 (17.4–31.2) | 30 | 20.8 (15.0–28.2) | 9 | 7 | 77.8 (45.3–93.7) | 7 | 77.8 (45.3–93.7) |
| >50 animals | 468 | 22 | 4.7 (3.1–7.1) | 75 | 16.0 (13.0–19.6) | 26 | 11 | 42.3 (25.5–61.1) | 17 | 65.4 (46.2–80.6) | |
| Human population density | Very low (<50 inh./km2) | 220 | 18 | 8.2 | |||||||
| Low (50–300 inh./km2) | 182 | 11 | 6 | ||||||||
| Semi-dense (300–1500 inh/km2) inh./km2) | 173 | 14 | 8.1 | ||||||||
| Dense (>1500 inh./km2) | 37 | 13 | 35.1 | ||||||||
| Köppen climate area * | Csa | 279 | 11 | 3.9 (2.2–6.9) | 25 | 9.0 (6.1–12.9) | 15 | 6 | 40.0 (19.8–64.3) | 8 | 53.3 (30.1–75.2) |
| Csb | 333 | 45 | 13.5 (10.2–17.6) | 80 | 24.0 (19.8–28.9) | 20 | 12 | 60.0 (38.7–78.1) | 16 | 80.0 (58.4–91.9) | |
| NUTSII Region | Norte | 259 | 37 | 14.3 (10.6–19.1) | 72 | 27.8 (22.7–33.6) | 15 | 9 | 60.0 (35.8–80.2) | 12 | 80.0 (54.8–93.0) |
| Centro | 148 | 9 | 6.1 (3.2–11.2) | 14 | 9.5 (5.7–15.3) | 9 | 4 | 44.4 (18.9–73.3) | 7 | 77.8 (45.3–93.7) | |
| Lisboa | 20 | 0 | 3 | 15.0 (5.2–36.0) | 1 | 0 | 0.0 (0.0–79.4) | 1 | 100 (20.7–100.0) | ||
| Alentejo | 185 | 10 | 5.4 (3.0–9.7) | 16 | 8.7 (5.4–13.6) | 10 | 5 | 50.0 (23.7–76.3) | 4 | 40.0 (16.8–68.7) | |
| Production system ** | Intensive | 246 | 34 | 13.8 (10.1–18.7) | 69 | 28.1 (22.8–33.0) | 15 | 9 | 60.0 (35.8–80.2) | 12 | 80.0 (54.8–93.0) |
| Extensive | 282 | 20 | 7.1 (4.6–10.7) | 23 | 8.2 (5.5–11.9) | 16 | 7 | 43.8 (23.1–66.8) | 8 | 50.0 (28.0–72.0) | |
| Production type ** | Dairy | 209 | 32 | 15.3 (11.1–20.8) | 56 | 26.8 (21.3–33.3) | 13 | 9 | 69.2 (42.4–87.3) | 10 | 76.9 (49.7–91.8) |
| Beef | 280 | 19 | 6.8 (4.4–10.4) | 32 | 11.4 (8.2–15.7) | 16 | 6 | 37.5 (18.5–61.4) | 9 | 56.3 (33.2–76.9) | |
| Fattening/Finishing | 39 | 3 | 7.7 (2.7–20.3) | 4 | 10.3 (4.1–23.6) | 2 | 1 | 50.0 (9.5–90.6) | 1 | 50.0 (9.5–90.6) | |
| Total | 612 | 56 | 9.2 (7.1–11.7) | 105 | 17.2 (14.4–20.4) | 35 | 18 | 51.4 (35.6–67.0) | 24 | 68.6 (52.0–81.5) | |
* Csa: hot summer Mediterranean climate; Csb: warm summer Mediterranean climate [24]; ** Data for production system and production type available for 528 animals in 31 farms.
Table 2.
Univariate GLMM results for Neospora caninum seroprevalence. Farm ID was included as a random effects factor.
Table 2.
Univariate GLMM results for Neospora caninum seroprevalence. Farm ID was included as a random effects factor.
| Risk Factors | Odds Ratio | Confidence Interval (95%) | p-Value |
|---|---|---|---|
| Human population density | |||
| Very low (<50 inhabitants/km2) | |||
| Low (50–300 inhabitants/km2) | |||
| Semi-dense (300–1500 inhabitants/km2) | 4.646 | 1.547–13.949 | 0.006 |
| Dense (>1500 inhabitants/km2) | 13.640 | 2.086–89.176 | 0.006 |
| Köppen climate area | |||
| Csa | |||
| Csb | 4.352 | 1.411–13.421 | 0.011 |
| NUTS2 region | |||
| Norte | 7.11 | 1.836–27.537 | 0.005 |
| Centro | |||
| Lisboa | |||
| Alentejo | |||
| Production system | |||
| Extensive | |||
| Intensive | 6.396 | 1.831–22.334 | 0.004 |
| Bioclimatic variable | |||
| bio13—Precipitation of wettest month | 1.024 | 1.007–1.041 | 0.006 |
| bio14—Precipitation of driest month | 1.154 | 1.037–1.283 | 0.009 |
| bio17—Precipitation of driest quarter | 1.032 | 1.001–1.057 | 0.01 |
| bio18—Precipitation of warmest quarter | 1.028 | 1.001–1.047 | 0.004 |
Table 3.
Univariate GLMM results for Toxoplasma gondii seroprevalence. Farm ID was included as a random effects factor.
Table 3.
Univariate GLMM results for Toxoplasma gondii seroprevalence. Farm ID was included as a random effects factor.
| Risk Factors | Odds Ratio | Confidence Interval (95%) | p-Value |
|---|---|---|---|
| Herd size | |||
| ≤50 | |||
| >50 | 0.126 | 0.042–0.377 | <0.001 |
| Population density (inhabitants/km2) | |||
| <50 | |||
| 50–300 | |||
| 300–1500 | |||
| >1500 | 15.786 | 1.682–148.139 | 0.0157 |
| Köppen climate area | |||
| Csa | |||
| Csb | 4.521 | 1.269–16.109 | 0.02 |
| Bioclimatic variables | |||
| bio13—Precipitation of wettest month | 1.026 | 1.007–1.045 | 0.006 |
| bio14—Precipitation of driest month | 1.149 | 1.022–1.292 | 0.02 |
| bio17—Precipitation of driest quarter | 1.031 | 1.004–1.058 | 0.025 |
| bio18—Precipitation of warmest quarter | 1.025 | 1.004–1.047 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Waap, H.; Bärwald, A.; Nunes, T.; Schares, G. Seroprevalence and Risk Factors for Toxoplasma gondii and Neospora caninum in Cattle in Portugal. Animals 2022, 12, 2080. https://doi.org/10.3390/ani12162080
AMA Style
Waap H, Bärwald A, Nunes T, Schares G. Seroprevalence and Risk Factors for Toxoplasma gondii and Neospora caninum in Cattle in Portugal. Animals. 2022; 12(16):2080. https://doi.org/10.3390/ani12162080
Chicago/Turabian StyleWaap, Helga, Andrea Bärwald, Telmo Nunes, and Gereon Schares. 2022. "Seroprevalence and Risk Factors for Toxoplasma gondii and Neospora caninum in Cattle in Portugal" Animals 12, no. 16: 2080. https://doi.org/10.3390/ani12162080
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.