
Exploring the Antibiotic Resistance Profile of Clinical Klebsiella pneumoniae Isolates in Portugal

 ,
,  ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Distribution of the K. pneumoniae Isolates
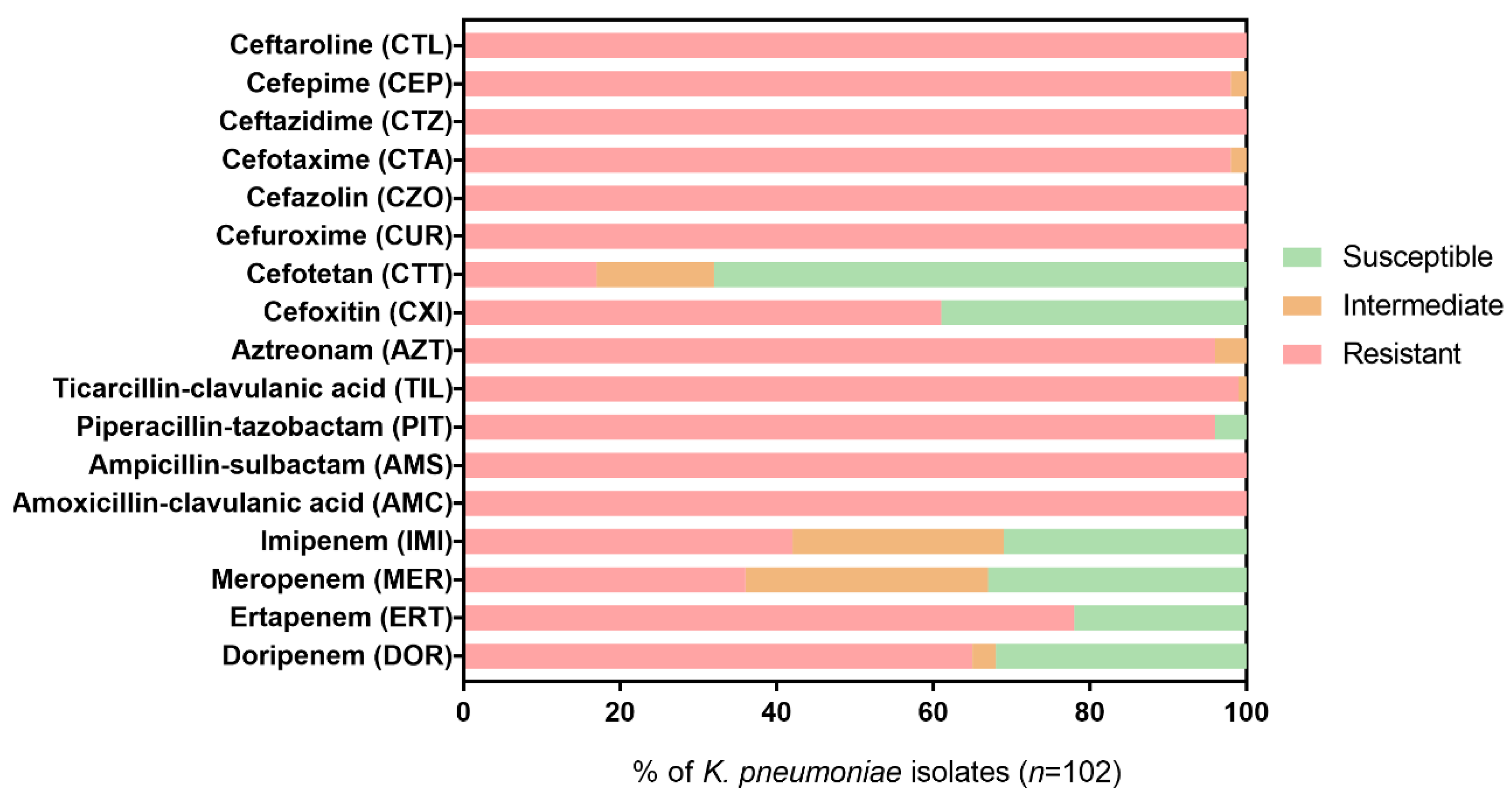
2.2. Resistance Profiles of K. pneumoniae Isolates to β-Lactam Antibiotics
2.3. Screening of Acquired β-Lactam Resistance Genes
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Susceptibility Testing
4.3. Detection of Acquired β-Lactam Resistance Genes by Multiplex PCR
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ballén, V.; Gabasa, Y.; Ratia, C.; Ortega, R.; Tejero, M.; Soto, S. Antibiotic Resistance and Virulence Profiles of Klebsiella pneumoniae Strains Isolated from Different Clinical Sources. Front. Cell Infect. Microbiol. 2021, 11, 824. [Google Scholar] [CrossRef] [PubMed]
- Blot, S.; Vandewoude, K.; Colardyn, F. Clinical Impact of Nosocomial Klebsiella Bacteremia in Critically Ill Patients. Eur. J. Clin. Microbiol. 2002, 21, 471–473. [Google Scholar] [CrossRef]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Genet. 2010, 8, 251–259. [Google Scholar] [CrossRef]
- Wei, J.; Wenjie, Y.; Ping, L.; Na, W.; Haixia, R.; Xuequn, Z. Antibiotic resistance of Klebsiella pneumoniae through β-arrestin recruitment-induced β-lactamase signaling pathway. Exp. Ther. Med. 2018, 15, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Ruppé, E.; Woerther, P.-L.; Barbier, F. Mechanisms of antimicrobial resistance in Gram-negative bacilli. Ann. Intensiv. Care 2015, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Toussaint, K.A.; Gallagher, J.C. β-Lactam/β-Lactamase Inhibitor Combinations: From Then to Now. Ann. Pharmacother. 2015, 49, 86–98. [Google Scholar] [CrossRef]
- Wanger, A.; Chavez, V.; Huang, R.S.P.; Wahed, A.; Actor, J.K.; Dasgupta, A. Antibiotics, Antimicrobial Resistance, Antibiotic Susceptibility Testing, and Therapeutic Drug Monitoring for Selected Drugs. In Microbiology and Molecular Diagnosis in Pathology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 119–153. [Google Scholar]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile β-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [Green Version]
- Cantón, R.; Akóva, M.; Carmeli, Y.; Giske, C.; Glupczynski, Y.; Gniadkowski, M.; Livermore, D.; Miriagou, V.; Naas, T.; Rossolini, G.; et al. Rapid evolution and spread of carbapenemases among Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 2012, 18, 413–431. [Google Scholar] [CrossRef] [Green Version]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine—3rd Revision; WHO: Geneva, Switzerland, 2011. [Google Scholar]
- Pitout, J.D.; Laupland, K.B. Extended-spectrum β-lactamase-producing Enterobacteriaceae: An emerging pub-lic-health concern. Lancet Infect. Dis. 2008, 8, 159–166. [Google Scholar] [CrossRef]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacoby, G.A. AmpC Β-Lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The European Committee on Antimicrobial Susceptibility Testing. EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. Version 2.0. 2017. Available online: http://www.eucast.org (accessed on 22 October 2022).
- Livermore, D.M. Bacterial Resistance: Origins, Epidemiology, and Impact. Clin. Infect. Dis. 2003, 36, S11–S23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Woodford, N.; Ellington, M.J.; Coelho, J.M.; Turton, J.F.; Ward, M.E.; Brown, S.; Amyes, S.G.; Livermore, D.M. Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrob. Agents 2006, 27, 351–353. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Gregson, D.B.; Poirel, L.; McClure, J.-A.; Le, P.; Church, D.L. Detection of Pseudomonas aeruginosa Producing Metallo-β-Lactamases in a Large Centralized Laboratory. J. Clin. Microbiol. 2005, 43, 3129–3135. [Google Scholar] [CrossRef] [Green Version]
- Mlynarcik, P.; Roderova, M.; Kolar, M. Primer Evaluation for PCR and its Application for Detection of Carbapenemases in Enterobacteriaceae. Jundishapur J. Microbiol. 2016, 9, e29314. [Google Scholar] [CrossRef] [Green Version]
- Arlet, G.; Rouveau, M.; Philippon, A. Substitution of alanine for aspartate at position 179 in the SHV-6 extended-spectrum β-lactamase. FEMS Microbiol. Lett. 1997, 152, 163–167. [Google Scholar] [CrossRef]
- Monstein, H.-J.; Östholm-Balkhed, A.; Nilsson, M.V.; Dornbusch, K.; Nilsson, L.E. Multiplex PCR amplification assay for the detection of blaSHV, blaTEM and blaCTX-M genes in Enterobacteriaceae. Apmis 2007, 115, 1400–1408. [Google Scholar] [CrossRef]
- Kojima, A.; Ishii, Y.; Ishihara, K.; Esaki, H.; Asai, T.; Oda, C.; Tamura, Y.; Takahashi, T.; Yamaguchi, K. Extended-Spectrum-β-Lactamase-Producing Escherichia coli Strains Isolated from Farm Animals from 1999 to 2002: Report from the Japanese Veterinary Antimicrobial Resistance Monitoring Program. Antimicrob. Agents Chemother. 2005, 49, 3533–3537. [Google Scholar] [CrossRef]
- World Health Organization. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug Resistant Bacterial Infections, Including Tuberculosis; World Health Organization: Geneva, Switzerland, 2017; pp. 1–88. [Google Scholar]
- Peterson, L.R. Bad bugs, no drugs: No ESCAPE revisited. Clin. Infect. Dis. 2009, 49, 992–993. [Google Scholar] [CrossRef] [PubMed]
- Bhagirath, A.Y.; Li, Y.; Patidar, R.; Yerex, K.; Ma, X.; Kumar, A.; Duan, K. Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens. Int. J. Mol. Sci. 2019, 20, 1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennequin, C.; Robin, F. Correlation between antimicrobial resistance and virulence in Klebsiella pneumoniae. Eur. J. Clin. Microbiol. 2015, 35, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, D.; Huang, X.; Long, S.; Yu, H. The Distribution of K. pneumoniae in Different Specimen Sources and Its Antibiotic Resistance Trends in Sichuan, China From 2017 to 2020. Front. Med. 2022, 9, 75214. [Google Scholar] [CrossRef] [PubMed]
- Miftode, I.-L.; Nastase, E.V.; Miftode, R.; Miftode, E.G.; Iancu, L.S.; Luncă, C.; Păduraru, D.-T.A.; Costache, I.-I.; Stafie, C.-S.; Dorneanu, O.-S. Insights into multidrug-resistant K. pneumoniae urinary tract infections: From susceptibility to mortality. Exp. Ther. Med. 2021, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Mangold, K.; Wyant, K.; Schora, D.M.; Voss, B.; Kaul, K.L.; Hayden, M.; Chundi, V.; Peterson, L.R. Rectal Screening for Klebsiella pneumoniae Carbapenemases: Comparison of Real-Time PCR and Culture Using Two Selective Screening Agar Plates. J. Clin. Microbiol. 2012, 50, 2596–2600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). The Evolving Threat of Antimicrobial Resistance: Options for Action; WHO Publications: Geneva, Switzerland, 2012; pp. 1–119. [Google Scholar]
- Lapi, F.; Marconi, E.; Pecchioli, S.; Lagolio, E.; Rossi, A.; Concia, E.; Cricelli, C. Reduced effectiveness among β-lactam antibiotics: A population-based cohort study in primary care in Italy. J. Antimicrob. Chemother. 2021, 76, 2186–2194. [Google Scholar] [CrossRef] [PubMed]
- Spadar, A.; Phelan, J.; Elias, R.; Modesto, A.; Caneiras, C.; Marques, C.; Lito, L.; Pinto, M.; Cavaco-Silva, P.; Ferreira, H.; et al. Genomic epidemiological analysis of Klebsiella pneumoniae from Portuguese hospitals reveals insights into circulating antimicrobial resistance. Sci. Rep. 2022, 12, 1–13. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control Antimicrobial Resistance in the EU/EEA (EARS-Net)—Annual Epidemiological Report for 2020. Available online: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-eueea-ears-net-annual-epidemiological-report-2020 (accessed on 2 November 2022).
- Meletis, G. Carbapenem resistance: Overview of the problem and future perspectives. Ther. Adv. Infect. Dis. 2015, 3, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Chastain, D.B.; White, B.P.; Cretella, D.A.; Bland, C.M. Is It Time to Rethink the Notion of Carbapenem-Sparing Therapy against Extended-Spectrum β-Lactamase–Producing Enterobacteriaceae Bloodstream Infections? Crit. Rev. Ann. Pharmacother. 2018, 52, 484–492. [Google Scholar] [CrossRef]
- Karaiskos, I.; Giamarellou, H. Carbapenem-sparing strategies for ESBL producers: When and how. Antibiotics 2020, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheikh, A.F.; Khoshnood, S.; Saki, M.; Shahi, F.; Seyed-Mohammadi, S.; Abdi, M.; Moradi, M. Prevalence of carbapenemases and ESBL encoding genes among K. pneumoniae isolates obtained from an educational hospital in Ahvaz, Southwestern Iran. Gene Rep. 2021, 23, 101128. [Google Scholar] [CrossRef]
- Bhaskar, B.H.; Mulki, S.S.; Joshi, S. Molecular Characterization of Extended Spectrum â-lactamase and Carbapenemase Producing Klebsiella pneumoniae from a Tertiary Care Hospital. Indian J. Crit. Care Med. 2019, 23, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Okanda, T.; Haque, A.; Koshikawa, T.; Islam, A.; Huda, Q.; Takemura, H.; Matsumoto, T.; Nakamura, S. Characteristics of Carbapenemase-Producing Klebsiella pneumoniae Isolated in the Intensive Care Unit of the Largest Tertiary Hospital in Bangladesh. Front. Microbiol. 2021, 11, 3357. [Google Scholar] [CrossRef] [PubMed]
- Desta, K.; Woldeamanuel, Y.; Azazh, A.; Mohammod, H.; Desalegn, D.; Shimelis, D.; Gulilat, D.; Lamisso, B.; Makonnen, E.; Worku, A.; et al. High Gastrointestinal Colonization Rate with Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae in Hospitalized Patients: Emergence of Carbapenemase-Producing K. pneumoniae in Ethiopia. PLoS ONE 2016, 11, e0161685. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.D.D.; Nordmann, P.; Poirel, L. Carbapenemase-Producing Klebsiella pneumoniae, a Key Pathogen Set for Global Nosocomial Dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef] [Green Version]
- Perdigão, J.; Caneiras, C.; Elias, R.; Modesto, A.; Spadar, A.; Phelan, J.; Campino, S.; Clark, T.G.; Costa, E.; Saavedra, M.J.; et al. Genomic Epidemiology of Carbapenemase Producing Klebsiella pneumoniae Strains at a Northern Portuguese Hospital Enables the Detection of a Misidentified Klebsiella variicola KPC-3 Producing Strain. Microorganisms 2020, 8, 1986. [Google Scholar] [CrossRef]
- Šuto, S.; Bedenić, B.; Likić, S.; Kibel, S.; Anušić, M.; Tičić, V.; Zarfel, G.; Grisold, A.; Barišić, I.; Vraneš, J. Diffusion of OXA-48 carbapenemase among urinary isolates of Klebsiella pneumoniae in non-hospitalized elderly patients. BMC Microbiol. 2022, 22, 1–15. [Google Scholar] [CrossRef]
- Kazmierczak, K.M.; Biedenbach, D.J.; Hackel, M.; Rabine, S.; de Jonge, B.L.M.; Bouchillon, S.K.; Sahm, D.F.; Bradford, P.A. Global dissemination of blaKPC into bacterial species beyond Klebsiella pneumoniae and in vitro susceptibility to ceftazidime-avibactam and aztreonam-avibactam. Antimicrob. Agents Chemother. 2016, 60, 4490–4500. [Google Scholar] [CrossRef] [Green Version]
- David, S.; Reuter, S.; Harris, S.R.; Glasner, C.; Feltwell, T.; Argimon, S.; Abudahab, K.; Goater, R.; Giani, T.; Errico, G.; et al. Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 2019, 4, 1919–1929. [Google Scholar] [CrossRef]
- Aires-De-Sousa, M.; de la Rosa, J.M.O.; Gonçalves, M.L.; Pereira, A.L.; Nordmann, P.; Poirel, L. Epidemiology of Carbapenemase-Producing Klebsiella pneumoniae in a Hospital, Portugal. Emerg. Infect. Dis. 2019, 25, 1632–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avgoulea, K.; Di Pilato, V.; Zarkotou, O.; Sennati, S.; Politi, L.; Cannatelli, A.; Themeli-Digalaki, K.; Giani, T.; Tsakris, A.; Rossolini, G.M.; et al. Characterization of Extensively Drug-Resistant or Pandrug-Resistant Sequence Type 147 and 101 OXA-48-Producing Klebsiella pneumoniae Causing Bloodstream Infections in Patients in an Intensive Care Unit. Antimicrob. Agents Chemother. 2018, 62, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guducuoglu, H.; Gursoy, N.C.; Yakupogullari, Y.; Parlak, M.; Karasin, G.; Sunnetcioglu, M.; Otlu, B. Hospital Outbreak of a Colistin-Resistant, NDM-1- and OXA-48-Producing Klebsiella pneumoniae: High Mortality from Pandrug Resistance. Microb. Drug Resist. 2018, 24, 966–972. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Carbapenemase-Producing (OXA-48) Klebsiella pneumoniae ST392 in Travellers Previously Hospitalised in Gran Canaria, Spain. 2018. Available online: https://www.ecdc.europa.eu (accessed on 22 October 2022).
- Rodrigues, C.; Novais, A.; Machado, E.; Peixe, L. Detection of VIM-34, a novel VIM-1 variant identified in the intercontinental ST15 Klebsiella pneumoniae clone. J. Antimicrob. Chemother. 2013, 69, 274–275. [Google Scholar] [CrossRef] [Green Version]
- Novais, Â.; Ferraz, R.V.; Viana, M.; da Costa, P.M.; Peixe, L. NDM-1 Introduction in Portugal through a ST11 KL105 Klebsiella pneumoniae Widespread in Europe. Antibiotics 2022, 11, 92. [Google Scholar] [CrossRef]
- Cantón, R.; Novais, A.; Valverde, A.; Machado, E.; Peixe, L.; Baquero, F.; Coque, T. Prevalence and spread of extended-spectrum β-lactamase-producing Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 2008, 14, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, C.; Machado, E.; Ramos, H.; Peixe, L.; Novais, Â. Expansion of ESBL-producing Klebsiella pneumoniae in hospitalized patients: A successful story of international clones (ST15, ST147, ST336) and epidemic plasmids (IncR, IncFIIK). Int. J. Med Microbiol. 2014, 304, 1100–1108. [Google Scholar] [CrossRef]
- Breurec, S.; Guessennd, N.; Timinouni, M.; Le, T.; Cao, V.; Ngandjio, A.; Randrianirina, F.; Thiberge, J.; Kinana, A.; Dufougeray, A.; et al. Klebsiella pneumoniae resistant to third-generation cephalosporins in five African and two Vietnamese major towns: Multiclonal population structure with two major international clonal groups, CG15 and CG258. Clin. Microbiol. Infect. 2013, 19, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Coque, T.M.; Baquero, F.; Canton, R. Increasing prevalence of ESBL-producing enterobacteriaceae in Europe. Eurosurveillance 2008, 13, 19044. [Google Scholar] [CrossRef]
- Lee, M.Y.; Ko, K.S.; Kang, C.-I.; Chung, D.R.; Peck, K.R.; Song, J.-H. High prevalence of CTX-M-15-producing Klebsiella pneumoniae isolates in Asian countries: Diverse clones and clonal dissemination. Int. J. Antimicrob. Agents 2011, 38, 160–163. [Google Scholar] [CrossRef]
- Woodford, N.; Reddy, S.; Fagan, E.J.; Hill, R.L.R.; Hopkins, K.L.; Kaufmann, M.E.; Kistler, J.; Palepou, M.-F.I.; Pike, R.; Ward, M.E.; et al. Wide geographic spread of diverse acquired AmpC -lactamases among Escherichia coli and Klebsiella spp. in the UK and Ireland. J. Antimicrob. Chemother. 2006, 59, 102–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulstad, C.R.; Solheim, M.; Berg, S.; Lindbæk, M.; Dahle, U.R.; Wester, A.L. Carriage of ESBL/AmpC-producing or ciprofloxacin non-susceptible Escherichia coli and Klebsiella spp. in healthy people in Norway. Antimicrob. Resist. Infect. Control 2016, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, F.; Machado, E.; Ribeiro, T.G.; Novais, Â.; Peixe, L. Long-term dissemination of acquired AmpC β-lactamases among Klebsiella spp. and Escherichia coli in Portuguese clinical settings. Eur. J. Clin. Microbiol. 2013, 33, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Sun, J.; Xu, X.; Huang, S. Epidemiology and risk factors of rectal colonization of carbapenemase-producing Enterobacteriaceae among high-risk patients from ICU and HSCT wards in a university hospital. Antimicrob. Resist. Infect. Control 2020, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Calderaro, A.; Buttrini, M.; Martinelli, M.; Montecchini, S.; Covan, S.; Ruggeri, A.; Rodighiero, I.; Di Maio, A.; Galullo, M.; Larini, S.; et al. Active surveillance for carbapenemase-producing Klebsiella pneumoniae and correlation with infection in subjects attending an Italian tertiary-care hospital: A 7-year retrospective study. BMJ Open 2021, 11, e042290. [Google Scholar] [CrossRef] [PubMed]
- Del Bianco, F.; Morotti, M.; Zannoli, S.; Dirani, G.; Fantini, M.; Pedna, M.F.; Farabegoli, P.; Sambri, V. Comparison of Four Commercial Screening Assays for the Detection of blaKPC, blaNDM, blaIMP, blaVIM, and blaOXA48 in Rectal Secretion Collected by Swabs. Microorganisms 2019, 7, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pages, J.-M.; Lavigne, J.-P.; Leflon-Guibout, V.; Marcon, E.; Bert, F.; Noussair, L.; Nicolas-Chanoine, M.-H. Efflux Pump, the Masked Side of ß-Lactam Resistance in Klebsiella pneumoniae Clinical Isolates. PLoS ONE 2009, 4, e4817. [Google Scholar] [CrossRef] [Green Version]
- Reyes, J.; Aguilar, A.C.; Caicedo, A. Carbapenem-resistant Klebsiella pneumoniae: Microbiology key points for clinical practice. Int. J. Gen. Med. 2019, 12, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Roschanski, N.; Fischer, J.; Guerra, B.; Roesler, U. Development of a Multiplex Real-Time PCR for the Rapid Detection of the Predominant Beta-Lactamase Genes CTX-M, SHV, TEM and CIT-Type AmpCs in Enterobacteriaceae. PLoS ONE 2014, 9, e100956. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- European Committe on Antimicrobial Susceptibility Testing. Antimicrobial Susceptibility Testing—EUCAST Disk Diffusion Version 10.0. 2022. Available online: http://www.eucast.org (accessed on 22 October 2022).
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: http://www.eucast.org (accessed on 22 October 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Category | Antimicrobial Agent (Abbreviation) |
|---|---|
| Carbapenems | Ertapenem (ERT) |
| Imipenem (IMI) | |
| Meropenem (MER) | |
| Doripenem (DOR) | |
| Penicillins + β-lactamase inhibitors | Amoxicillin-clavulanic acid (AMC) |
| Ampicillin-sulbactam (AMS) | |
| Antipseudomonal penicillins + β-lactamase inhibitors | Ticarcillin-clavulanic acid (TIL) |
| Piperacillin-tazobactam (PIT) | |
| Monobactams | Aztreonam (AZT) |
| Cephamycins | Cefoxitin (CXI) |
| Cefotetan (CTT) | |
| Non-extended-spectrum cephalosporins; 1st and 2nd generation cephalosporins | Cefazolin (CZO) |
| Cefuroxime (CUR) | |
| Extended-spectrum cephalosporins; 3rd and 4th generation cephalosporins | Cefotaxime (CTA) |
| Ceftazidime (CTZ) | |
| Cefepime (CEP) | |
| Anti-MRSA cephalosporins | Ceftaroline (CTL) |
| Target Gene | Primer (5’–3’) | Amplicon Size | Annealing Temperature | Ref. | |
|---|---|---|---|---|---|
| Multiplex 1 Class A and class D carbapenemases | blaKPC-like | CGTCTAGTTCTGCTGTCTTG | 798 bp | 60 °C | [17] |
| CTTGTCATCCTTGTTAGGCG | |||||
| blaOXA-48-like | TTGGTGGCATCGATTATCGG | 743 bp | [17] | ||
| GAGCACTTCTTTTGTGATGGC | |||||
| blaOXA-23-like | GATCGGATTGGAGAACCAGA | 501 bp | [18] | ||
| ATTTCTGACCGCATTTCCAT | |||||
| Multiplex 2 Class B carbapenemases (MBL) | blaIMP-like | GAAGGYGTTTATGTTCATAC | 587 bp | 54 °C | [19] |
| GTAMGTTTCAAGAGTGATGC | |||||
| blaNDM-1-like | TAAAATACCTTGAGCGGGC | 439 bp | [20] | ||
| AAATGGAAACTGGCGACC | |||||
| blaVIM-like | GTTTGGTCGCATATCGCAAC | 382 bp | [19] | ||
| AATGCGCAGCACCAGGATAG | |||||
| Multiplex 3 ESBL | blaTEM-like | CATTTYCGTGTCGCCCTTATTC | 800 bp | 60 °C | [13] |
| CGTTCATCCATAGTTGCCTGAC | |||||
| blaSHV-like | AGCCGCTTGAGCAAATTAAAC | 713 bp | [21] | ||
| ATCCCGCAGATAAATCACCAC | |||||
| blaCTX-M-like | ATGTGCAGYACCAGTAARGTKATGGC | 593 bp | [22] | ||
| TGGGTRAARTARGTSACCAGAAYCAGCGG | |||||
| Multiplex 4 AmpC β-lactamases | blaCMY-2-like | ATGATGAAAAAATCGTTATGCT | 1145 bp | 60 °C | [23] |
| TTATTGCAGCTTTTCAAGAATGCG | |||||
| blaDHA-like | TGATGGCACAGCAGGATATTC | 997 | [13] | ||
| GCTTTGACTCTTTCGGTATTCG | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, R.; Castro, J.; Silva, S.; Oliveira, H.; Saavedra, M.J.; Azevedo, N.F.; Almeida, C. Exploring the Antibiotic Resistance Profile of Clinical Klebsiella pneumoniae Isolates in Portugal. Antibiotics 2022, 11, 1613. https://doi.org/10.3390/antibiotics11111613
Oliveira R, Castro J, Silva S, Oliveira H, Saavedra MJ, Azevedo NF, Almeida C. Exploring the Antibiotic Resistance Profile of Clinical Klebsiella pneumoniae Isolates in Portugal. Antibiotics. 2022; 11(11):1613. https://doi.org/10.3390/antibiotics11111613
Chicago/Turabian StyleOliveira, Ricardo, Joana Castro, Sónia Silva, Hugo Oliveira, Maria José Saavedra, Nuno Filipe Azevedo, and Carina Almeida. 2022. "Exploring the Antibiotic Resistance Profile of Clinical Klebsiella pneumoniae Isolates in Portugal" Antibiotics 11, no. 11: 1613. https://doi.org/10.3390/antibiotics11111613